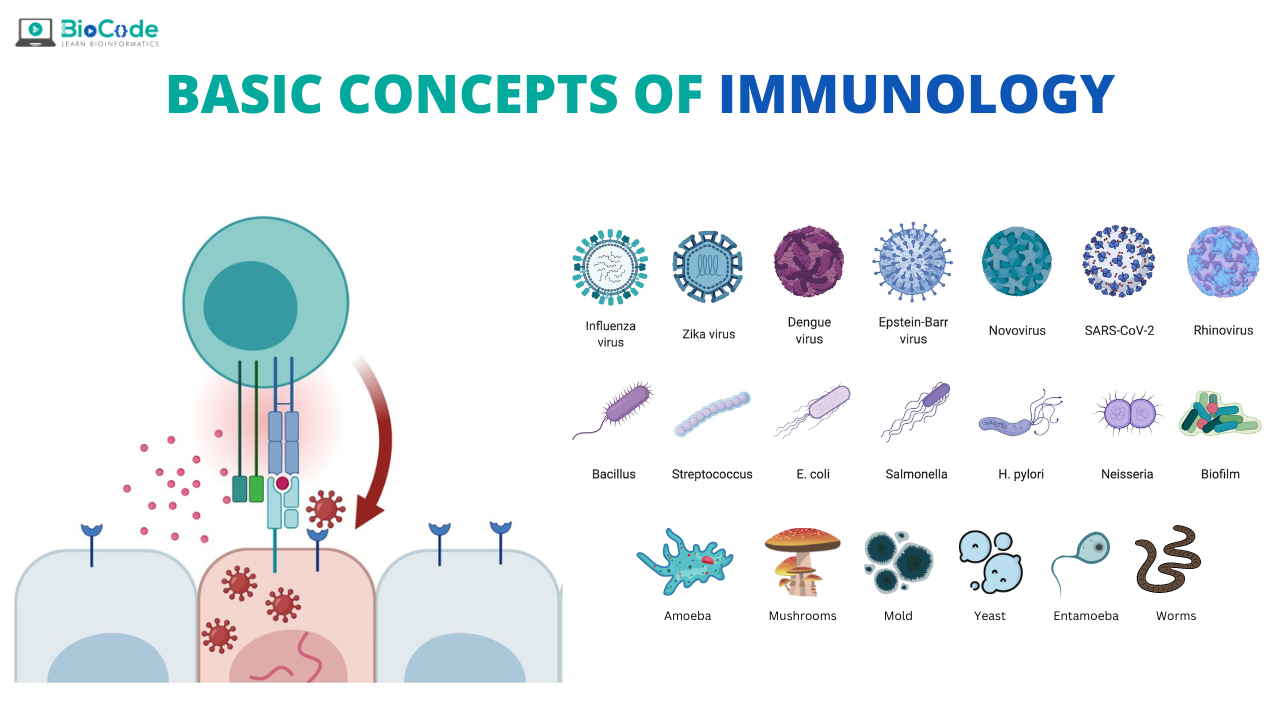
Pathogen
An organism that can cause diseases are known as pathogens.
Pathogenesis:
The process of inducing disease in an organism is called as pathogenesis. Examples of different pathogens representing major categories are
Bacteria: E. Coli cause food borne illness.
Virus: Influenza virus cause flu.
Fungi: Aspergillus flavus produces cancerous aflatoxin.
Immune Barriers:
Barriers are needed to be breached for immune system activation.
Barriers includes skin, mucous membrane, acidity of stomach and vagina, perspiration, and internal defense e. g complement proteins. An animal can break this barrier by biting which can expose the inside to the pathogens for infections and diseases. For example Rabies, Tetanus, Malaria and Plagues.
Pathogen Recognition:
The process of pathogen recognition involves an interaction between the foreign organism and a recognition molecule expressed by host cells. Host cell can express membrane bound receptor, solid receptor and secretory recognition molecules. While ligands for these molecules can be whole pathogen, fragments or secreted products. Changes in the host cell after being infected is detected by T lymphocytic cells and cytotoxic T cells. T cells binds to the virus or viral proteins in cytosol and kill the virus infected cell. The killing of cell can sometimes affect the surrounding normal cells for example in HIV, the T helper cells are infected which disrupt the balance of immune response. B cells also help in destroying pathogens. The antibodies produced can reach places which are not accessible to the B cells. Parasites, which are too big for phagocytes to capture, they are treated by the cells which can deposit toxic substances or induce expulsion like sneezing and coughing.
Most pathogens express at least a few chemical structures that are not typically found in mammals. Pathogen-associated molecular patterns (or PAMPs) are common foreign structures that characterize whole groups of pathogens. It is these unique antigenic structures that the immune system frequently recognizes first. Animals, both invertebrates and vertebrates, have evolved to express several types of cell surface and soluble proteins that quickly recognize many of these PAMPs: a form of pathogen profiling. White blood cells naturally express a variety of receptors, collectively referred to as patterns recognition receptors (PRRs), that specifically recognize these sugar residues, as well as other common foreign substances. When PRRs detect these chemical structures, a cascade of events labels the target pathogen for destruction. PRRs are proteins encoded in the genomic DNA and are always expressed by many different immune cells. These conserved, germline-encoded recognition molecules are thus a first line of defense for the quick detection of many of the typical chemical identifiers carried by the most common invaders. Bacteria have LPS, DNA, Flagellin. Viruses have viral DNA, and its coat proteins and Parasites have GPI anchors.
Generation of diversity:
This phenomenon happens in developing B and T cells which give rise to a group of B and T cells where each expresses many copies of one unique recognition molecule, resulting in a population with the theoretical potential to respond to any antigen that may come along. This is accomplished by rearranging and editing the genomic DNA that encodes the antigen receptors expressed by each B or T lymphocyte. Not unlike the error-prone DNA replication method employed by pathogens, this system allows chance to play a role in generating different recognition molecules.
Maturation in T and B cells, which occurs in primary lymphoid organs (bone marrow for B cells and thymus for T cells) in the absence of antigen, produces cells with a committed antigenic specificity, each of which expresses many copies of surface receptor that binds to one antigen. Cells that do not die or become deleted during this maturation and weeding-out process move into the circulation of the body and are available to interact with antigen. There, clonal selection occurs when one of these cells encounters its cognate or specific antigen. Clonal proliferation of an antigen-activated cell leads to many cells that can engage with and destroy the antigen, plus memory cells that can be called upon during a subsequent exposure. The B cells secrete antibody, a soluble form of the receptor, reactive with the activating antigen. Similar processes take place in the T-lymphocyte population, resulting in clones of memory T cells and effector T cells; the latter include activated TH cells, which secrete cytokines that aid in the further development of adaptive immunity, and cytotoxic T lymphocytes (CTLs), which can kill infected host cells.
Tolerance
The self and non-self-discrimination relies on the principle of tolerance. Burnet (Nobel prize winner) was the first to propose that exposure to non-self-antigens during certain stages of life could result in an immune system that ignore these antigens in later life. Experiment was performed on mouse embryo for this theory. To establish tolerance, the antigen receptors present on developing B and T cells must first pass a test of non-responsiveness against host structures.Most recently another hypothesis is proposed called “danger hypothesis”. According to this hypothesis, the immune system constantly evaluates each new encounter more for its potential to be dangerous to the host than for whether it is self or not. Dysfunctional tolerance is at the root of most autoimmune diseases.
Innate and adaptive Immunity:
There are two interconnected systems of immunity: innate and adaptive. These two systems collaborate to protect the body against foreign invaders. Innate immunity includes built-in molecular and cellular mechanisms that are encoded in the germ line and are evolutionarily more primitive, aimed at preventing infection or quickly eliminating common invaders. A second form of immunity, known as adaptive immunity, is much more attuned to subtle molecular differences. This part of the system, which relies on B and T lymphocytes, takes longer to come on board but is much more antigen specific.
Primary & Secondary Immune Response & Memory Generation:
When an animal is injected with an antigen, it produces a primary antibody response of low magnitude and short duration, peaking at about 10 to 20 days. At some later point, a second exposure to the same antigen results in a secondary response that is greater in magnitude. It peaks in less time (1–4 days) and is more antigen specific than the primary response. Innate responses, which have no memory element and occur each time an antigen is encountered, are unchanged regardless of how frequently this antigen has been encountered in the past.
Innate And Adaptive Immune Response to An Infection:
Pathogens are introduced at a mucosal surface or breach in skin, where they are picked up by phagocytic cells. In innate stage of the response, the phagocytic cell undergoes changes and carries pieces of bacteria to a lymph node to help activate adaptive immunity. Meanwhile, at the site of infection resident phagocytes encountering antigen release chemokines and cytokines that cause fluid influx and help recruit other immune cells to the site (inflammation). In the lymph node, T and B cells with appropriate receptor specificity are clonally selected when their surface receptors bind antigen that has entered the system, kicking off adaptive immunity. Collaboration between T and B cells and continued antigen encounter occurs in the lymph node, driving lymphocyte proliferation and differentiation, generating cells that can very specifically identify and eradicate the pathogen. For example: B cells secrete antibodies specific for the antigen, which travel to the site of infection to help label and eradicate the pathogen. In addition to the cells that will destroy the pathogen here, memory T and B cells are generated in this primary response and will be available at the initiation of a secondary response, which will be much more rapid and antigen specific.
